
Yehu Moran
Feel free to contact Yehu: yehu.moran at mail.huji.ac.il / yehum79 at yahoo.com
Yehu studied from 2001-2010 at Tel Aviv University where he obtained a BSc in Life Sciences, MSc in Biochemistry and PhD working on sea anemone toxins. Then he moved to the lab of Ulrich Technau at the University of Vienna to study the evolution of post-transcriptional regulation by small RNAs. At January 2014 he was appointed as a senior lecturer at the Department of Ecology, Evolution and Behavior of the Alexander Silberman Institute of Life Sciences of The Hebrew University of Jerusalem and nowadays he is a Professor at this department. He is currently serving as an elected Department head. (picture below by Yonit Schiller and the Hebrew University).

Our Research
I am an experimental biologist. I find the interface between bioinformatics, genomics and experimental biology very exciting but I'm not a CS person. I was trained in a wide range of biochemical techniques as my master degree is in biochemistry. In my PhD my topics of research switched more into evolution and in my postdoc I also added developmental biology into the mix. Later as a faculty at Jerusalem I also became fascinated by organismal biology and molecular and chemical ecology. I believe cnidarians are a wonderful research system if you want to get a perspective about what animal life looked like 600 million years ago since this is when they diverged from the rest of the animals. Despite being considered "primitive" they share with us humans sophisticated cell types, including neurons and muscles. The interesting topics you can study with this system are endless. The thread that connects all our research topics is the evolution of complex systems. We wish to understand how extremely complex system are formed and maintained across large evolutionary timescales despite their apparent cost. You can find below our current research topics:
Research Topics


The evolution of Innate immunity and Antiviral systems in Animals
Viruses are everywhere. They are absolute parasites and there is no organism that does not have a virus that can infect it. Viruses are a major force in evolution and they are locked in an “arm-race” with the immune systems of their host cells. We started in May 2020 a new ERC Consolidator project called AntiViralEvo. In this project we study the evolution of antiviral systems in cnidarians, with emphasis on Anthozoa (sea anemones and corals) and their RNA—based detection and response systems. As an initial step we characterized the virome of our favorite anthozoan, Nematostella. Our first functional study of the cnidarian antiviral system was published in 2021 in MBE (see here). It unravels the evolution of the RIG-I Like Receptors (RLRs) and their function in Nematostella. You can also find popular science coverage (in Hebrew) here. Addtionally, we started in May 2021 a collaborative project (funded by BSF-NSF joint grants) with the lab of Adam Reitzel at the University of North Carolina at Charlotte where we compare the viromes and antiviral systems of different Nematostella populations to understand their evolution under ecological context.
The evolution of small RNA pathways and the roles of microRNAs in Cnidaria
MicroRNAs are small RNAs which were discovered initially in nematodes at 1993. A decade later they were also identified in most other animals and in plants and were shown to play crucial roles in the development of many species. MicroRNAs act through a protein effector complex which binds to messenger RNAs in a specific manner via annealing of the microRNA to the mRNA target and affect the target expression levels. I initialized during my postdoc in Vienna a study on the roles of microRNAs in N. vectensis development. This study is still underway but we already obtained some very exciting findings regarding the involvement of microRNAs in the regulation of expression levels of proteins involved in development of the anemone. Moreover, we have interesting results regarding the mechanism of action of microRNAs in cnidarians suggesting new and unexpected links between the microRNA pathways of plants and animals see our papers in MBE and Genome Research about this topic. This project also received much hype (and somewhat disinformative presentation!) in popular media such as here, here (in English) and here (in Hebrew). You can also read our review in Nature Ecology and Evolution here. We published in 2018 the first evidence for the role of microRNAs and piRNAs in Nematostella development and the role of methylation in stabilizing these small RNAs (see here). In 2020 we published a paper in Nature Communications deciphering the specialization of Argonaute proteins in sea anemones and corals in carrying different classes of miRNAs and siRNAs and unravelling how miRNAs are born from siRNAs in Cnidaria (see here). In 2022 we published in eLife our findings regarding a cnidarian homolog of the “plant-specific” HYL1 protein and its role in miRNA biogenesis, which puts in question the traditional evolutionary scenarios of the miRNA pathway (see here). Our latest work on miRNAs focused on the pan-cnidarian miRNA miR-2022 and revealed its function in the biogenesis of cnidocytes (the famous cnidarian stinging cells; see here).
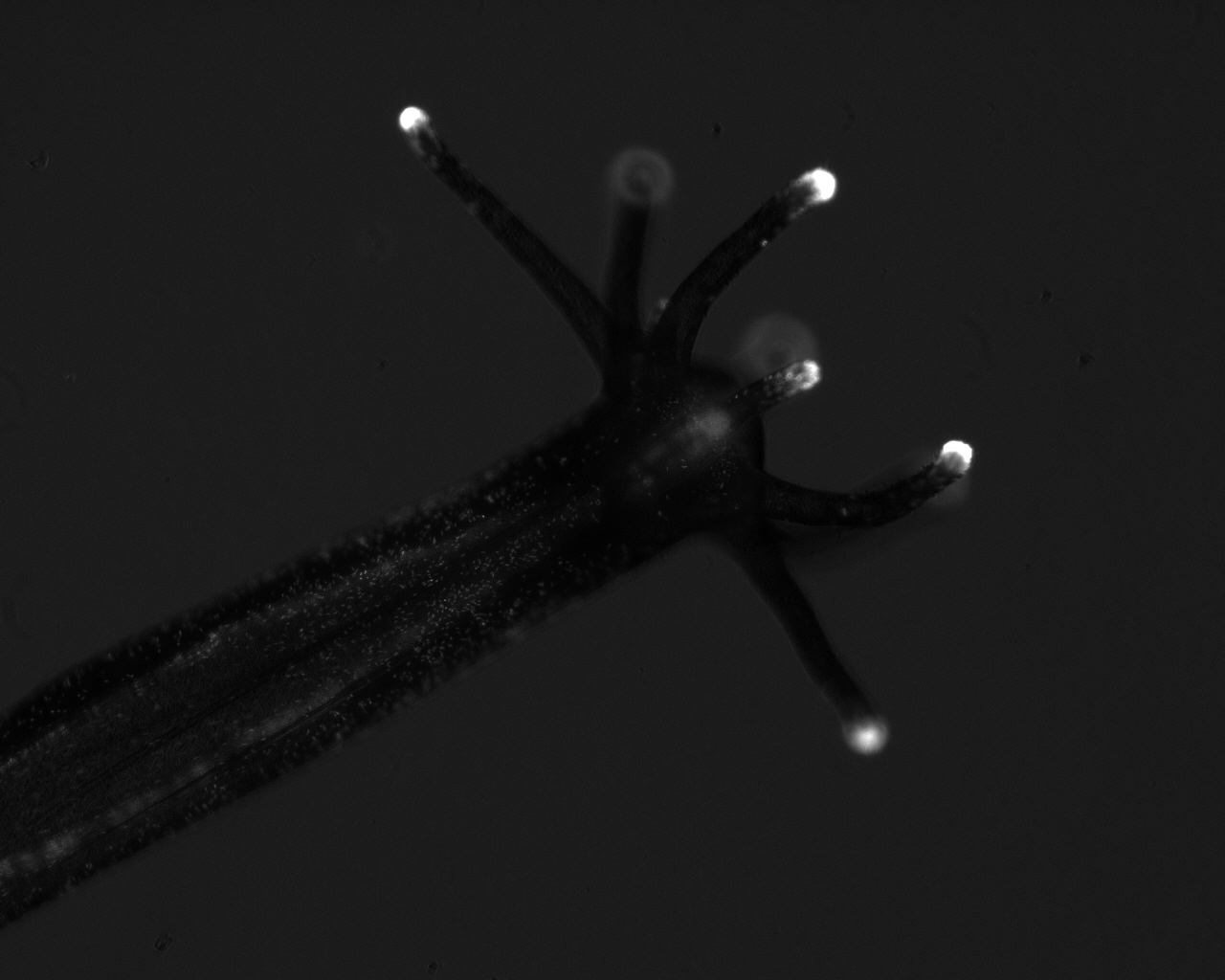
Evolution of sea anemone (and other strange animal) toxins
This is a study I initialized as a PhD student in Tel Aviv University at the lab of Prof. Michael Gurevitz and I pursued it further in my postdoc as a side project (toxinology is a very sad addiction) and now as a PI. We published several papers about the unusual evolutionary patterns we discovered in sea anemone and scorpion toxins and speculated regarding the factors driving their selection. In general, animal toxins just like immunology-related proteins are a vast playground for strong adaptive selection as they are involved in a never-ending "arms-race" of prey and predator. We also found evidence for an unusual mode of evolution, called "concerted evolution" in the toxin genes of sea anemones. Further, we also show that the expression of neurotoxins happens in several anemone species such as Nematostella not only in nematocytes (stinging cells) but in gland cells (see here). This finding challenges the common statement presented in many invertebrate zoology textbooks that peptide cnidarian toxins are produced exclusively in nematocytes. The ecological shifts which led to toxin expression in different cell populations of closely-related species are of great interest and open the field to eco-evo studies. We also employed proteomic methods for the study of the venom components of Nematosella nematocytes and published those results.
We are taking advantage on unique tools such as transgenesis techniques that are available in Nematostella for answering questions about the evolution of venom and venom-producing cells. You can see recent examples here, here and here. You can find popular science pieces about our work from Haaretz news paper here (in Hebrew) and here (in English).
We are also fascinated by the effect of venom on organismal fitness and the ecology of animals. This is a new major direction that our lab is taking. See our latest work here and the news piece (in English) from the Jerusalem Post here.

Evolution of ION channels
Voltage-gated sodium channels are pivotal components in the conductance of neuronal signals. The unprecedented extent of sequencing of animal genomes in the last few years has enabled us to study in detail the evolution of these channels. We found that these channels have first appeared in unicellular organisms before the split of fungi and animals, but were independently lost in many lineages. Surprisingly cnidarians have a remarkable diversity of channel isoforms, but most of them cannot discern sodium from calcium and potassium. However, Cnidarians do have a selective channel isoform that evolved more than 540 million years ago in the ancestor of all extant cnidarians. The molecular basis of this selectivity is different from that found in selective sodium channels of bilaterians, indicating that sodium selectivity evolved twice independently in animals, probably in order to comply with the rising need for more complex and faster neuronal transmission. In this work we applied both electrophysiological and phylogenetic methods and a first publication came out in Cell Reports. It was followed by a review paper. Nowadays we also study the function of the DEG/ENaC channels of Nematostella. Members of this channel superfamily exhibit an extraordinary level of functional and gating diversity, leading us to wonder and investigate what might have been their original function in basally-branching animals. See our paper on the evolution of this channel superfamily and its role in sea anemones here.

Funding
Our research is generously funded by the Binational Science Foundation (BSF), the European Research Council (ERC), German-Israeli Foundation for Scientific Research and Development (GIF), the Israel Council of Higher Learning, the Israel Science Foundation (ISF) and the Marie Curie Actions of the European Commission. We are grateful for this support.
LAB MEMBERS

Reuven Aharoni
(Lab manager) reuven.Aharoni at mail.huji.ac.il
Rubi obtained his MSc in Biomedical Studies and his PhD from the Institute of Dental Sciences of the Hebrew University. Before joining our lab in 2014 and taking important part in its establishment, he was working in the lab of Doron Steinberg on bacterial biofilm. He is our lab manager and is responsible for everything running smoothly. Rubi significantly contributed to our understanding of the evolution of Argonaute proteins and their function in Cnidaria. Recently, he was taking part in our functional studies of cnidarian innate immunity.

Yael Admoni (PhD student) yael.admoni at mail.huji.ac.il
Yael studied towards her BSc in Life Sciences in the Etgar program for excellent biology students at the Hebrew University. During her 3rd and final year she performed in our lab her final student science project. From 2017 until 2019 she was a master student in the lab working on knocking out specific miRNA genes in Nematostella and on the complementarity requirements of cnidarian miRNA targets. Yael rejoined our group as a PhD student in late 2020 after working for several months as a reseaech assistant. Her PhD work is focusing on revealing the roles of RNA interference in cnidarian biology and the evolution of this ancient system.

Daria Aleshkina (Postdoc) daria.aleshkina at mail.huji.ac.il
Daria completed her Ph.D. project in the laboratory of Dr. Andrej Susor at the Institute of Animal Physiology and Genetics, Czech Academy of Science, and at Charles University, Prague. She focused on the roles of non-coding RNAs during the meiotic maturation of mammalian oocyte and early embryo development. Daria joined our laboratory in September 2023 as a postdoc and studies the RNAi pathway as a part of the innate immunity of Nematostella vectensis, with a particular focus on the RNA-dependent RNA polymerases (RdRPs). She intends to discover in which molecular aspects of antiviral response RdRPs are involved in N. vectensis.

Yael Hazan (PhD student) yael.hzn at gmail.com
Yael received her BSc in Biology from the Hebrew university and her MSc in Marine Biology from Ben Gurion University. She started her PhD in our lab in April 2017 studying the evolution of RNA-based immunity mechanisms in Cnidaria. One of her main research focuses is revealing the roles of Cnidarian RdRPs (RNA-dependent RNA Polymerases). Yael’s projects include a significant computational component and she is co-advised by Prof. Michal Linial, a computational biologist from the Department of Chemical Biology at our University.

Adrian Jaimes-Becerra (postdoc) ajjb80 at gmail.com
Adrian joined our lab as a postdoc on October 2021. Before joining our group he obtained his PhD from São Paulo University (Brazil). He worked there with Prof. Antonio Carlos Marques on the molecular evolution and phylogeny of cnidarian toxins. In our group Adrian works on the functional genetics of Nematostella venom, applying genetic-manipulation tools for revealing the roles of specific venom components.

Itamar Kozlovski (PhD student) itamar.kozlovski at mail.huji.ac.il
Itamar majored in Biology at Colby-Sawyer College (New Hampshire, USA) and obtained his MSc from the Hebrew University Medical School in 2017. During his MSc Itamar studied under the guidance of Prof. Rotem Karni the roles of RNA splicing in cancer biology. Since May 2020 he is a PhD student in our group and studies the antiviral response of Nematostella. Itamar integrates into his work advanced cell biology methods and fluorescent-activated cell sorting.

Hee-Jin Kwak (postdoc) someting2 at gmail.com
Hee-Jin obtained his MSc and PhD in 2021 from the Program of Zoology and Conservation Biology of Chungbuk National University (South Korea). He was working in his PhD on evo-devo of leeches in the lab of Prof. Sung-Jin Cho. He joined our group in November 2021 and his current research focus is the role of OAS1 and RNAse L in the cnidarian innate immunity as well as evolutionary comparisons of the antiviral innate immune systems of different Nematostella populations.

Ton Sharoni (PhD student) sharoni.rko at gmail.com
Ton obtained his BSc in Marine Biotechnology from the Ruppin Academic Center (Michmoret Campus). He joined our lab in October 2017 as an MSc student to study the function of RLR proteins in Nematostella. In December 2019 he graduated and rejoined the lab in April 2020 as a PhD student. His current project deals with the genetic manipulation of RLRs and their partner proteins as well as expanding into the relationship between stress and antiviral immunity in Cnidaria.

Joachim M. Surm (postdoc) joachim.surm at mail.huji.ac.il
Dr. Joachim Surm received his PhD with distinction from the Queensland University of Technology (QUT) in Brisbane, Australia At QUT Joachim worked on the molecular evolution of toxins and other proteins in sea anemones. He joined our group in February 2019 and he studies the evolution of toxins and toxin-like proteins in Nematostella as well as their variation between populations. He is also currently working on genetic mamipulation of venom genes in order to understand their contribution to organismal fitness. Joachim received the Golda Meir and Lady Davis Fellowships from the Hebrew University.
Lab Alumni

Jose Maria Aguilar Camacho (postdoc 2019-2021). Now postdoc at Chrstina Zakas’ lab at North Carolina State University, Raleigh, USA.

Yaara Y. Columbus-Shenkar (PhD student 2015-2019). Now lab manager of Matan Golan’s lab, ARO Volcani.

Arie Fridrich (PhD student and postdoc 2015-2020; postdoc 2021). Now postdoc at Frederic Berger’s group, GMI, Vienna, Austria.

Magda Lewandowska (postdoc 2018-2020; senior postdoc 2020-2021). Now bioinformatician at Ardigen.

Vengamanaidu Modepalli (Postdoc 2015-2018). Now Anne Warner Fellow at the Marine Biology Association, Plymouth, UK.

Maria Y. Sachkova (postdoc 2015-2018). Now postdoc at Pawel Burkhardt’s lab, Sars Centre, Bergen, Norway.

Kartik Sunagar (postdoc and Marie Curie Fellow 2014-2017). Now Assistant Professor at the Indian Institute of Science, Banagalor, India.

Abhinandan Mani Tripathi (postdoc 2019-2021).

Yoav Tal (MSc student 2014-2016).

Shir Singer (MSc student 2016-2018).

Morani Landau (MSc student 2018-2021).
Publications
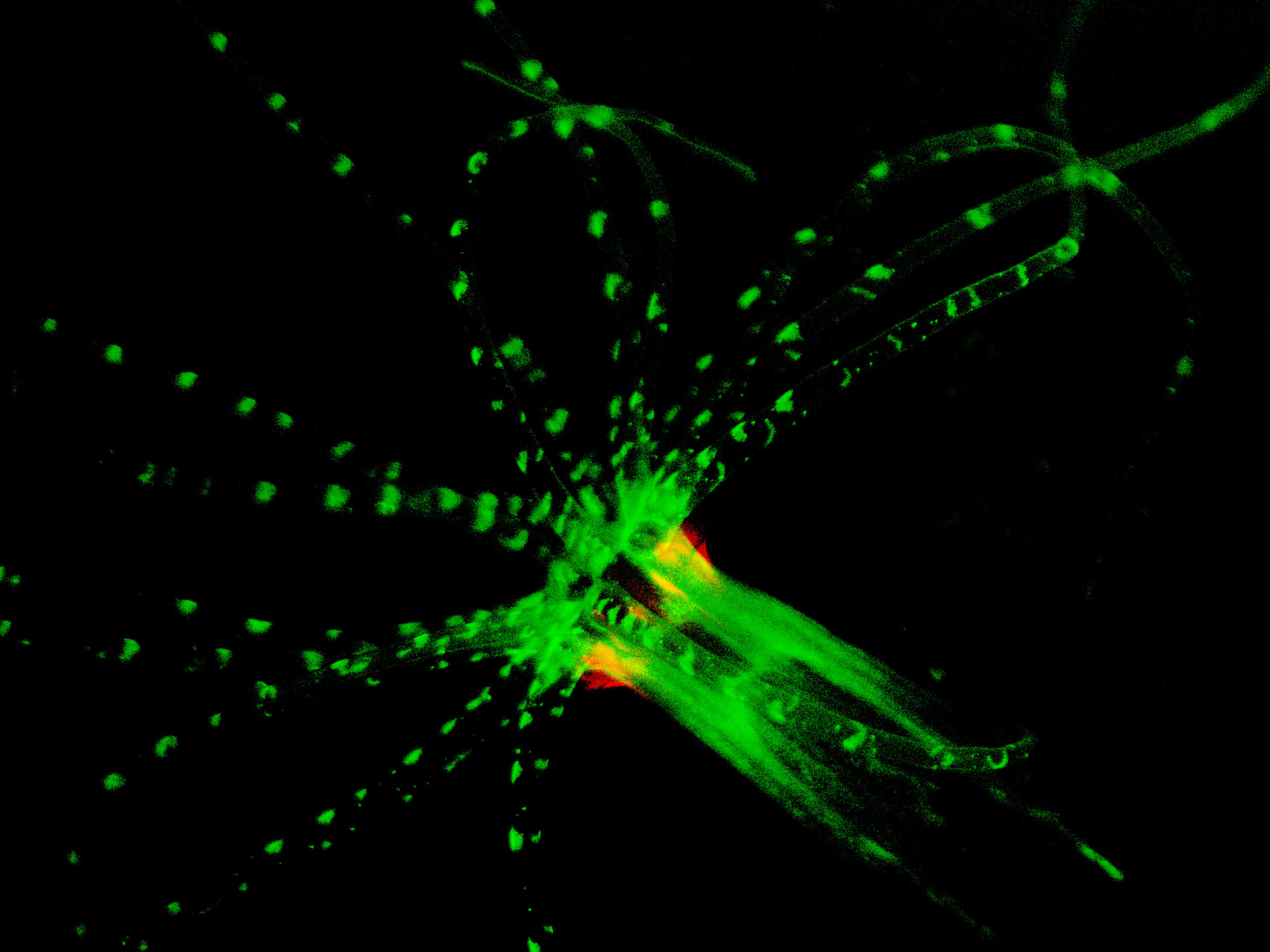
Research Papers
Key: *equal contribution; #corresponding author(s)
53. Sharoni T, Jaimes-Becerra A, Lewandowska M, Aharoni R, Voolstra CR, Fine M, Moran Y# (2025) Heat Stress Drives Rapid Viral and Antiviral Innate Immunity Activation in Hexacorallia. Molecular Ecology 34: e70098 link
52. Rutlekowski AI#, Modepalli V, Ketchum R, Moran Y, Reitzel AM.De novo genome assembly of the Edwardsiid anthozoan Edwardsia elegans. G3 15: jkaf011 link
51. Admoni Y#, Fridrich A, Weavers PK, Aharoni R, Razin T, Salinas-Saavedra M, Rabani M, Frank U, Moran Y# (2025) Target complementarity in cnidarians supports a common origin for animal and plant microRNAs. EMBO Reports Accepted. doi: 10.1038/s44319-024-00350-z link
50. Talice S, Kozlovski I, Barkan SK, Snyder GA, Sharoni T, Levy T, Oisher S, Ottolenghi A, Eliachar S, Ben-Romano R, Berlyne K, Yannai R, Lewandowska M, Sultan E, Goldstein O, Aharoni R, Hadad U, Davis C, Moran Y, Gershoni-Yahalom O, Traylor-Knowles N#, Rosental B# (2024) Candidate stem cell isolation and transplantation in Hexacorallia. Cell Reports 43: 114944 link
49. Deneke VE, Blaha A, Lu Y, Suwita JP, Draper JM, Phan CS, Panser K, Schleiffer A, Jacob L, Humer T, Stejskal K, Krssakova G, Roitinger E, Handler D, Kamoshita M, Vance TDR, Wang X, Surm JM, Moran Y, Lee JE, Ikawa M, Pauli A# (2024) A conserved fertilization complex bridges sperm and egg in vertebrates. Cell 10: S0092-8674(24)01093-6 link
48. Kozlovski I#*, Jaimes-Becerra A*, Sharoni T, Lewandowska M, Karmi O, Moran Y# (2024) Induction of apoptosis by double-stranded RNA was present in the last common ancestor of cnidarian and bilaterian animals. PLOS Pathogens 20: e1012320 link
47. Surm JM#, Landau M, Columbus-Shenkar YY, Moran Y# (2024) Sea anemone MACPF proteins demonstrate an evolutionary transitional state between venomous and developmental functions. Mol. Biol. Evol. 41: msae082 link
46. Surm JM#, Birch S, Macrander J, Jaimes-Becerra A, Fridrich A, Aharoni R, Rozenblat R, Sharabany J, Appelbaum L, Reitzel AM, Moran Y# (2024) Venom trade-off shapes interspecific interactions, physiology, and reproduction. Sci. Adv. 10: eadk3870 link
45. Zimmermann B*, Montenegro JD*, Robb SMC, Fropf WJ, Weilguny L, He S, Chen S, Lovegrove-Walsh J, Hill EM, Chen CY, Ragkousi K, Praher D, Fredman D, Schultz D, Moran Y, Simakov O, Genikhovich G, Gibson MC#, Technau U# (2023) Topological structures and syntenic conservation in sea anemone genomes. Nat. Commun. 14: 8270 link
44. Fridrich A#, Salinas-Saaverda M, Kozlolvski I, Surm JM, Chrysostomou E, Tripathi AM, Frank U, Moran Y# (2023) An ancient pan-cnidarian microRNA regulates stinging capsule biogenesis in Nematostella vectensis. Cell Rep. 42: 113072 link
43. Gert KRB, Panser K, Surm J, Steinmetz BS, Schleiffer A, Jovine L, Moran Y, Kondrashov F, Pauli A# (2023) Divergent molecular signatures in fish Bouncer proteins define cross-fertilization boundaries. Nat. Commun. 14: 3506 link
42. Smith EG*#, Surm JM*#, Macrander J, Simhi A, Amir G, Sachkova MY, Lewandowska M, Reitzel AM, Moran Y# (2023) Micro and macroevolution of sea anemone venom phenotype. Nat. Commun. 14: 249 link
41. Aguilar-Camacho JM*, Foreman K*, Jaimes-Becerra A, Aharoni R, Gründer S#, Moran Y# (2023) Functional analysis in a model sea anemone reveals phylogenetic complexity and a role in cnidocyte discharge of DEG/ENaC ion channels. Commun. Biol. 6: 17 link
40. Eliachar S, Snyder GA, Barkan SK, Talice S, Otolenghi A, Jaimes-Becerra A, Sharoni T, Sultan E, Hadad U, Levy O, Moran Y, Gershoni-Yahalom O, Traylor-Knowles N#, Rosental B#. (2022) Heat stress increases immune cell function in Hexacorallia. Front. Immunol. 13: 1016097 link
39. Senji Laxme RR, Khochare S, Attarde S, Kaur N, Jaikumar P, Shaikh NY, Aharoni R, Primor N, Hawlena D, Moran Y#, Sunagar K# (2022) The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii. Toxins 14: 725. link
38. Taiber S, Gozlan O, Cohen R, Andrade LR, Gregory EF, Starr DA, Moran Y, Hipp R, Kelley MW, Manor U, Sprinzak D#, Avraham KB# (2022) A Nesprin-4/kinesin-1 cargo model for nuclear positioning in cochlear outer hair cells Front. Cell Dev. Biol. 10: 974168 link
37. Tripathi AM*, Admoni Y*, Fridrich A, Lewandowska M, Surm JM, Aharoni R, Moran Y# (2022) Functional characterization of a “plant-like” HYL1 homolog in the cnidarian Nematostella vectensis indicates a conserved involvement in microRNA biogenesis. eLife 11: e69464 link
36. Lewandowska M#, Sharoni T, Admoni Y, Aharoni R, Moran Y# (2021) Functional characterization of the cnidarian antiviral immune response reveals ancestral complexity. Mol. Biol. Evol. 38: 4546-4561 link
35. Praher D, Zimmermann B, Dnyansagar R, Miller DJ, Moya A, Modepalli V, Fridrich A, Sher D, Friis-Møller L, Sundberg P, Fôret S, Ashby R, Moran Y#, Technau U# (2021) Conservation and turnover of miRNAs and their highly complementary targets in early branching animals. Proc. R. Soc. B 288: 20203169. link
34. Fridrich A*, Modepalli V*, Lewandowska M, Aharoni R, Moran Y# (2020) Unravelling the developmental and functional significance of an ancient Argonaute duplication. Nat. Commun. 11: 6187. *equal contribution link
33. Sachkova MY#, Landau M*, Surm JM*, Macrander J, Singer SA, Reitzel AM, Moran Y# (2020) Toxin-like neuropeptides in the sea anemone Nematostella unravel recruitment from the nervous system to venom. Proc. Natl. Acad. Sci. U.S.A 117: 27481-27492. *equal contribution pubmed link
32. Admoni Y*, Kozlovski I*, Lewandowska M, Moran Y# (2020) TATA Binding Protein (TBP) Promoter Drives Ubiquitous Expression of Marker Transgene in the Adult Sea Anemone Nematostella vectensis. Genes (Basel) 11: E1081. *equal contribution pubmed link
31. Sachkova MY#, Macrander J, Surm JM, Aharoni R, Menard-Harvey SS, Klock A, Leach WB, Reitzel AM#, Moran Y# (2020) Some like it hot: population-specific adaptations in venom production to abiotic stressors in a widely distributed cnidarian. BMC Biol. 18: 121 pubmed link
30. Tournière O, Dolan D, Richards GS, Sunagar K, Columbus-Shenkar YY, Moran Y, Rentzsch F# (2020) NvPOU4/Brain3 Functions as a Terminal Selector Gene in the Nervous System of the Cnidarian Nematostella Vectensis. Cell Rep. 30: 4473-4489. pubmed link
29. Lewandowska M#, Hazan Y, Moran Y# (2020) Initial Virome Characterization of the Common Cnidarian Lab Model Nematostella vectensis. Viruses 12: 218. pubmed link
28. Sachkova MY*#, Singer SA*, Macrander J, Reitzel AM, Peigneur S, Tytgat J, Moran Y# (2019) The birth and death of toxins with distinct functions: a case study in the sea anemone Nematostella. Mol. Biol. Evol. 36: 2001-2012. pubmed link
27. Sunagar K*#, Columbus-Shenkar YY*, Fridrich A, Gutkovitch N, Aharoni R, Moran Y# (2018) Cell type-specific expression profiling unravels the development and evolution of stinging cells in sea anemone. BMC Biol. 16: 108. pubmed link
26. Modepalli V*#, Fridrich A*, Agron M*, Moran Y# (2018) The methyltransferase HEN1 is required in Nematostella vectensis for microRNA and piRNA stability as well as larval metamorphosis. PLOS Genet. 14: e1007590. *equal contribution. pubmed link
25. Dnyansagar R, Zimmermann B, Moran Y, Praher D, Sundberg P, Møller LF, Technau U# (2018) Dispersal and speciation: The cross Atlantic relationship of two parasitic cnidarians. Mol. Phylogenet. Evol. 126: 346-355. pubmed link
24. Columbus-Shenkar YY*, Sachkova MY*, Macrander J, Fridrich A, Modepalli V, Reitzel AM, Sunagar K, Moran Y# (2018) Dynamics of venom composition across a complex life cycle. eLife 7: e35014. pubmed link
23. Praher D, Zimmermann B, Genikhovich G, Columbus-Shenkar Y, Modepalli V, Aharoni R, Moran Y#, Technau U# (2017) Characterization of the piRNA pathway during development of the sea anemone Nematostella vectensis. RNA Biol. 14: 1727-1741. pubmed link
22. Modepalli V#, Moran Y# (2017) Evolution of miRNA Tailing by 3' Terminal Uridylyl Transferases in Metazoa. Genome Biol. Evol. 9: 1547-1560. pubmed link
21. Mauri M, Kirchner M, Aharoni R, Ciolli Mattioli C, van den Bruck D, Gutkovitch N, Modepalli V, Selbach M, Moran Y, Chekulaeva M# (2017) Conservation of miRNA-mediated silencing mechanisms across 600 million years of animal evolution. Nucleic Acids Res. 45: 938-950 pubmed link
20. Sunagar K#, Moran Y# (2015) The Rise and Fall of an Evolutionary Innovation: Contrasting Strategies of Venom Evolution in Ancient and Young Animals. PLOS Genet. 11: e1005596. Link.
19. Jouiaei M*, Sunagar K*, Gross AF, Scheib H, Alewood PF, Moran Y#, Fry BG# (2015) Evolution of an ancient venom: recognition of a novel family of cnidarian toxins and the common evolutionary origin of sodium and potassium neurotoxins in sea anemone. Mol. Biol. Evol. 32:1598-610. pubmed link
18. Moran Y#, Zakon HH# (2014) The evolution of the four subunits of voltage-gated calcium channels: ancient roots, increasing complexity and multiple losses. Genome Biol. Evol. 6: 2210-7. pubmed link
17. Gur Barzilai M, Kahn R, Regev N, Gordon D, Moran Y, Gurevitz M# (2014) The specificity of Av3 sea anemone toxin for arthropods is determined at linker DI/SS2-S6 in the pore module of target sodium channels. Biochem. J. 463: 271-7. pubmed link
16. Moran Y, Fredman D, Praher D, Li XZ, Wee LM, Rentzsch F, Zamore PD#, Technau U#, Seitz H# (2014) Cnidarian microRNAs frequently regulate targets by cleavage. Genome Res. 24: 651-63. pubmed link
15. Moran Y#, Praher D, Fredman D, Technau U (2013) The evolution of microRNA pathway protein components in Cnidaria. Mol. Biol. Evol. 30: 2541-52. pubmed link
14. Orts DJ, Moran Y, Cologna CT, Peigneur S, Madio B, Praher D, Quinton L, De Pauw E, Bicudo JE, Tytgat J, de Freitas JC# (2013) BcsTx3 is a founder of a novel sea anemone toxin family of potassium channel blockers. FEBS J. 280: 4839-52. pubmed link
13. Nesher N#, Shapira E, Sher D, Moran Y, Tsveyer L, Turchetti-Maia AL, Horowitz M, Hochner B, Zlotkin E (2013) AdE-1, a new inotropic Na+ channel toxin from Aiptasia diaphana, is similar to, yet distinct from, known anemone Na+ channel toxins. Biochem. J. 451: 81-90. pubmed link
12. Moran Y#, Praher D, Schlesinger A, Ayalon A, Tal Y, Technau U (2013) Analysis of soluble protein contents from the nematocysts of a model sea anemone sheds light on venom evolution. Mar. Biotechnol. (NY). 15: 329-39. pubmed link
11. Gur Barzilai M, Reitzel AM, Kraus JEM, Gordon D, Technau U, Gurevitz M, Moran Y# (2012) Convergent evolution of sodium ion selectivity in metazoan neuronal signaling. Cell Rep. 2: 242-248. pubmed link
10. Moran Y*#, Fredman D*, Szczesny P, Grynberg M, Technau U# (2012) Recurrent horizontal transfer of bacterial toxin genes to eukaryotes. Mol. Biol. Evol. 29: 2223-2230. *equal contribution. pubmed link
9. Moran Y#, Genikhovich G, Gordon D, Wienkoop S, Zenkert C, Özbek S, Technau U, Gurevitz M (2012) Neurotoxin localization to ectodermal gland cells uncovers an alternative mechanism of venom delivery in sea anemones. Proc. R. Soc. B 279: 1351-1358. pubmed link
8. Weinberger H*, Moran Y*,Gordon D, Turkov M, Kahn R, Gurevitz M (2010) Positions under positive selection – key for selectivity and potency of scorpion alpha-toxins. Mol. Biol. Evol. 27: 1025-34. *these authors contributed equally to this work. pubmed link
7. Moran Y, Weinberger H, Lazarus N, Gur M, Kahn R, Gordon D, Gurevitz M (2009) Fusion and retrotransposition events in the evolution of sea anemone neurotoxin genes. J. Mol. Evol. 69: 115-24. pubmed link
6. Cohen L*, Moran Y*, Sharon A, Segal D, Gordon D, Gurevitz M (2009) An innate immunity peptide of Drosophila melanogaster, interacts with the fly voltage-gated sodium channel. J. Biol. Chem. 284: 23558-63. *these authors contributed equally to this work. pubmed link
5. Moran Y, Weinberger H, Reitzel AM, Sullivan JC, Kahn R, Gordon D, Finnerty JR, Gurevitz, M (2008) Intron retention as a post-transcriptional regulatory mechanism of neurotoxin expression at early life stages of the starlet anemone Nematostella vectensis. J. Mol. Biol. 380: 437-43. pubmed link
4. Moran Y, Weinberger H, Sullivan JC, Reitzel AM, Finnerty JR, Gurevitz M (2008) Concerted Evolution of sea anemone neurotoxin genes is revealed through analysis of the Nematostella vectensis genome. Mol. Biol. Evol. 25: 737-47. pubmed link
3. Moran Y, Kahn R, Cohen L, Gur M, Karbat I, Gordon D, Gurevitz M (2007) Molecular analysis of the sea anemone toxin Av3 reveals selectivity to insects and demonstrates the heterogeneity of receptor site-3 on voltage-gated Na-channels. Biochem. J. 406: 41-48. pubmed link
2. Moran Y, Gurevitz M (2006) When positive selection of neurotoxin genes is missing: The riddle of the sea anemone Nematostella vectensis. FEBS J. 273: 3886-92. pubmed link
1. Moran Y, Cohen L, Kahn R, Karbat I, Gordon D, Gurevitz M (2006) Mutagenesis of the sea anemone toxin Av2 reveals key amino acid residues important for activity on insect voltage-gated sodium channels. Biochemistry 45: 8864-73. pubmed link
REVIEW, Opinion and Re-Analysis ARTICLES (all peer reviewed)
14. Zancolli G#, von Reumont BM, Anderluh G, Caliskan F, Chiusano ML, Fröhlich J, Hapeshi E, Hempel BF, Ikonomopoulou MP, Jungo F, Marchot P, de Farias TM, Modica MV, Moran Y, Nalbantsoy A, Procházka J, Tarallo A, Tonello F, Vitorino R, Zammit ML, Antunes A# (2024) Web of venom: exploration of big data resources in animal toxin research. Gigascience 13: giae054 link
13. Fridrich A#, Moran Y# (2023) Some flies do not play ping-pong. PLOS Biol. 21: e3002152. link
12. Iwama RE#, Moran Y# (2023) Origins and diversification of animal innate immune responses against viral infections Nat. Eco. Evol. 7: 182-193 link
11. von Reumont BM#, Anderluh G, Antunes A, Ayvazyan N, Beis D, Caliskan F, Crnković A, Damm M, Dutertre S, Ellgaard L, Gajski G, German H, Halassy B, Hempel BF, Hucho T, Igci N, Ikonomopoulou MP, Karbat I, Klapa MI, Koludarov I, Kool J, Lüddecke T, Ben Mansour R, Vittoria Modica M, Moran Y, Nalbantsoy A, Ibáñez MEP, Panagiotopoulos A, Reuveny E, Céspedes JS, Sombke A, Surm JM, Undheim EAB, Verdes A, Zancolli G (2022) Modern venomics-Current insights, novel methods, and future perspectives in biological and applied animal venom research. Gigascience 11: giac048 link
10. Surm JM#, Moran Y# (2021) Transposons Increase Transcriptional Complexity: The Good Parasite? Trends Genet 37: 606-607. pubmed link
9. Modica MV#, Ahmad R, Ainsworth S, Anderluh G, Antunes A, Beis D, Caliskan F, Serra MD, Dutertre S, Moran Y, Nalbantsoy A, Oukkache N, Pekar S, Remm M, von Reumont BM, Sarigiannis Y, Tarallo A, Tytgat J, Undheim EAB, Utkin Y, Verdes A, Violette A, Zancolli G (2021) The new COST Action European Venom Network (EUVEN)-synergy and future perspectives of modern venomics. GigaScience 10: giab019. link
8. Surm JM#, Moran Y# (2021) Insights into how development and life-history dynamics shape the evolution of venom. EvoDevo 12: 1. link
7. Ul-Hasan S#, Rodríguez-Román E, Reitzel AM, Adams RM, Herzig V, Trim SA, Saviola AJ, Nobile CJ, Stiers EE, Moschos SA, Keiser CN, Petras D, Moran Y, Colston TJ# (2019) The emerging field of venom-microbiomics for exploring venom as a microenvironment, and the corresponding Initiative for Venom Associated Microbes and Parasites (iVAMP). Toxicon: X 10016. link
6. Fridrich A#, Hazan Y, Moran Y# (2019) Too Many False Targets for MicroRNAs: Challenges and Pitfalls in Prediction of miRNA Targets and Their Gene Ontology in Model and Non-model Organisms. Bioessays 41: e1800169. pubmed link
5. Moran Y#, Agron M, Praher D, Technau U# (2017) The evolutionary Origin of plant and animal microRNAs. Nat. Ecol. Evol. 1: 0027. pubmed link
4. Sunagar K, Morgenstern D#, Reitzel AM, Moran Y# (2015) Ecological venomics: How genomics, transcriptomics and proteomics can shed new light on the ecology and evolution of venom. J. Proteomics S1874-3919: 30131-7. pubmed link
3. Moran Y#, Gur Barzilai M, Liebeskind BJ, Zakon HH# (2015) Evolution of voltage-gated ion channels at the emergence of Metazoa. J. Exp. Biol. 218: 515-525. pubmed link
2. Qiu X, Brown, KV, Moran Y, Chen D# (2010) Sirtuin regulation in calorie restriction. Biochim. Biophys. Acta, 1804: 1576-83. pubmed link
1. Moran Y#, Gordon D, Gurevitz M# (2009) Sea anemone toxins affecting voltage-gated sodium channels - molecular and evolutionary features. Toxicon, 54: 1089-1101. pubmed link
Preprints (currently under review or revision)
3. Kozlovski I#, Jaimes-Becerra A, Aleshkina D, Levy M, Moran Y# (2025) Epigenomic and transcriptomic analyses reveal cnidocyte specialization in a sea anemone. bioRxiv doi: https://doi.org/10.1101/2025.10.06.680633
2. Kozlovski I#, Sharoni T, Levy S, Jaimes-Becerra A, Talice S, Kwak H, Aleshkina D, Aharoni R, Grau-Bové X, Karmi O, Rosental B, Sebe-Pedros A, Moran Y# (2025) Functional characterization of specialized immune cells in a cnidarian reveals an ancestral antiviral program. bioRxiv doi: https://doi.org/10.1101/2025.01.24.634691
1. Admoni Y#, Lewandowska M, Aharoni R, Shi J, Zhang X, Chen Q, Moran Y# (2025) Characterization of RNA interference in the model cnidarian Nematostella vectensis reveals partial target silencing but lack of small RNA amplification. bioRxiv doi: https://doi.org/10.1101/2025.01.03.631050


OuR Address
Yehu Moran's lab Room 3-344, Silberman building Department of Ecology, Evolution and Behavior
Alexander Silberman Institute of Life Sciences Faculty of Science Edmond J. Safra campus, Givat Ram The Hebrew University of Jerusalem 9190401 Jerusalem Israel
Email: yehu.moran at mail.huji.ac.il OR yehum79 at yahoo.com

The Hebrew University of Jerusalem
Lab pictures

Anemone system




Arie and Yael Maria and the FPLC Yaara and a crocodile


The view of western Jerusalem from our former lab Lab excursion to the Jerusalem zoo 2016

Moran lab group picture, May 2019

Moran lab group picture, August 2020

Moran lab group picture, December 2021